
Introduction
Man is the creature that can learn to learn.
I recently linked Five Articles on Anxiety, the first article being a paper my girlfriend Jennifer Kokkines wrote. After reading my girlfriend’s paper I did some research, linked in the article I wrote, but continued to ponder. I could find few to no papers which presented research solely on anxiety and its effects on decision making. Sure, I could find papers on fear, but that is not the same as anxiety. It astounds me that research has not been done (or at least not research I could discover) on this topic, its ripe for the picking! Therefore, I decided to dedicate this article to my girlfriend’s paper, which I felt was very insightful and at least brought to the table a discussion Neurology and Psychology researchers need to be having. Research into this topic would have implications in the field(s) of neurology, psychology, economics, marketing, sales, business, and more. Most importantly (for me anyways) it would have implications on how I (or you) learn and react. I began this article with a rather poorly phrased statement:
Man is the creature that can learn to learn.
Humans are the only creatures (I know of) which can learn about themselves to improve themselves. This is one of our greatest assets, we can take insights we gain about ourselves (good or bad) and use those insights to gain more knowledge. This would be called a “closed-loop” in control theory:
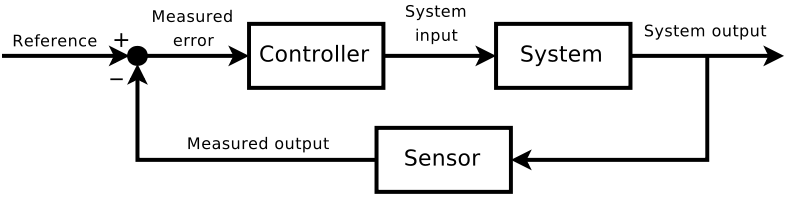
Humans can constantly take sensory inputs about ourselves to further improve ourselves. This is why the first steps to self-improvement must be understand what effects your decision-making. Hence the reason this is such an important area of research, and why I am frustrated that there is not enough research out there to quench my thirst to understand/learn.
I would also like to apologize that much of the research in this article is behind a paywall.
Anxiety and its Effect on Decision-Making
Jennifer Kokkines
Senior, Undergraduate
University of Illinois Urbana-Champaign
The dictionary definition of anxiety according to the Merriam-Webster dictionary):
A painful or apprehensive uneasiness of mind usually over an impending or anticipated ill
or
An abnormal and overwhelming sense of apprehension and fear often marked by physiological signs (as sweating, tension, and increased pulse), by doubt concerning the reality and nature of the threat, and by self-doubt about one’s capacity to cope with it.
In short, anxiety is essentially an inner state of turmoil an individual experiences during scenarios that don’t necessarily require such an intense response. Often, people who experience high anxiety will display certain behavioral traits, such as pacing back and forth, suffer from physical pain, or ruminate (focus on the symptoms of their own distress and the causes/consequences of it).
After conducting some research and delving into the world of the highly anxious, there seems to be strong evidence supporting that anxiety and the behavior it creates have some dictation over decision-making. The integration of neuroeconomics and the neural circuitry of fear learning play a large role in developing anxious traits and utilizing them to make specific choices.
Fear vs Anxiety
Understanding the difference between fear and anxiety is crucial in interpreting the data gathered from many studies and is also significant because both impact the other in the aspect of cognitive function. First off, as described in an article published at the National Institute of Health:
Fear responses are elicited by specific stimuli, and tend to be short-lived, decreasing once a threat has dissipated. Anxiety may be experienced in the absence of a direct physical threat, and typically persists over a long period of time. [1]
That being said, though the two share similar physiological properties, they both can be differentiated considering the way they respond and their reasons for their responses are not derived from the same root cause. Even the responses themselves differ based on length of time experienced.
The Hardware
It is speculated that the one of the main components in supporting fear conditioning in animal and human models is the amygdala, which plays a central role in fear acquisition, storage, and expression. As noted in one article published by Cisler:
A threat detection mechanism likely underlies facilitated attention, a process that may be neurally centered around the amygdala. Attentional control ability likely underlies difficulty in disengagement, emotion regulation goals likely underlie attentional avoidance, and both of these processes may be neurally centered around prefrontal cortex functioning. The threat detection mechanism may be a mostly automatic process, attentional avoidance may be a mostly strategic process, and difficulty in disengagement may be a mixture of automatic and strategic processing.[2]
It is quite possibly the designated location in which association and memory storage can cue fear conditioning and project to the brainstem and hypothalamus, and also project to the ventral striatum. The brainstem and the hypothalamus mediate autonomic fear expression, while the ventral striatum mediates the utilization of actions in order to cope with fear. Along with fear acquisition, the hippocampus also plays a vital role in modulating our fear. To boil it all down, the amygdala, ventromedial prefrontal cortex, and the hippocampus all contribute to the acquisition, storage, retrieval and contextual modulation of fear acquisition and extinction. Extinction is when conditioned responding to a stimulus decreases once the reinforcer has been removed. 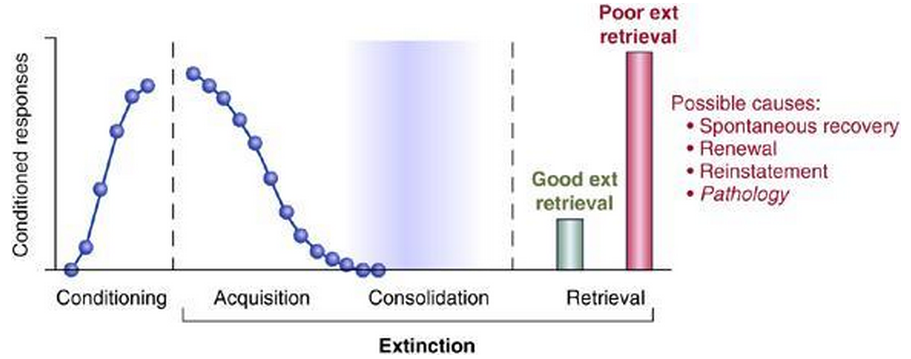 Caption for the image above: Extinction learning occurs in three phases. Acquisition is characterized by a decrease in conditioned responses to the presentation of a CS without the US. Consolidation is a time-dependent process during which a long-term extinction representation is formed. Retrieval of extinction occurs at a later time, when the CS is re-presented. Good extinction retrieval is characterized by low levels of conditioned responses (green bar), whereas poor extinction retrieval is characterized by high levels of conditioned responses (red bar). Poor retrieval of extinction is normally observed following renewal, reinstatement, spontaneous recovery, or in pathological conditions characterized by extinction failure.[3] In the duration of this extinction retrieval, projections from the ventromedial prefrontal cortex direct towards the amygdala’s inhibitory interneurons and diminish fear expression. Once extinction occurs, the contextual information will modulate competition between the original fear memory and the new extinction memory. During intentional fear regulation, activation of the amygdala typically decreases due to dorsolateral prefrontal cortex activation. This, in turn, activates the ventromedial prefrontal cortex-amygdala inhibitory pathway that mediates extinction rezrieval. One study that was done by Shmuel Lissek performed a review “representing fear-learning scores for 453 anxiety patients and 455 healthy controls, was conducted to . . . assess the moderating influences of study characteristics.” The results of this study revealed increases in acquisition of fear learning and conditioned responding during extinction in patients with anxiety. “ . . . These patient-control differences . . . primarily emerge from studies employing simple, single-cue paradigms where only danger cues are presented and no inhibition of fear to safety cues is required.” [4] Interestingly enough, there are other areas of the brain that can be correlated to and may contribute to the emotional expression and awareness that is associated with anxiety. For instance, a person who expresses anxiety is noted to have a heightened perception of bodily sensations – this is also referred to as “interoception”. Interoception may increase the aversiveness of a threat response. It is suggested that the insula plays a critical role in representing the interoceptive information. This is crucial, considering altered insula activity could skew modulation of an individual’s interoceptive awareness, thus contributing to such anxiety.[5]
Caption for the image above: Extinction learning occurs in three phases. Acquisition is characterized by a decrease in conditioned responses to the presentation of a CS without the US. Consolidation is a time-dependent process during which a long-term extinction representation is formed. Retrieval of extinction occurs at a later time, when the CS is re-presented. Good extinction retrieval is characterized by low levels of conditioned responses (green bar), whereas poor extinction retrieval is characterized by high levels of conditioned responses (red bar). Poor retrieval of extinction is normally observed following renewal, reinstatement, spontaneous recovery, or in pathological conditions characterized by extinction failure.[3] In the duration of this extinction retrieval, projections from the ventromedial prefrontal cortex direct towards the amygdala’s inhibitory interneurons and diminish fear expression. Once extinction occurs, the contextual information will modulate competition between the original fear memory and the new extinction memory. During intentional fear regulation, activation of the amygdala typically decreases due to dorsolateral prefrontal cortex activation. This, in turn, activates the ventromedial prefrontal cortex-amygdala inhibitory pathway that mediates extinction rezrieval. One study that was done by Shmuel Lissek performed a review “representing fear-learning scores for 453 anxiety patients and 455 healthy controls, was conducted to . . . assess the moderating influences of study characteristics.” The results of this study revealed increases in acquisition of fear learning and conditioned responding during extinction in patients with anxiety. “ . . . These patient-control differences . . . primarily emerge from studies employing simple, single-cue paradigms where only danger cues are presented and no inhibition of fear to safety cues is required.” [4] Interestingly enough, there are other areas of the brain that can be correlated to and may contribute to the emotional expression and awareness that is associated with anxiety. For instance, a person who expresses anxiety is noted to have a heightened perception of bodily sensations – this is also referred to as “interoception”. Interoception may increase the aversiveness of a threat response. It is suggested that the insula plays a critical role in representing the interoceptive information. This is crucial, considering altered insula activity could skew modulation of an individual’s interoceptive awareness, thus contributing to such anxiety.[5]
Trait Anxiety
Dysregulation of this neurocircuitry is proposed to have a role in the maintenance of anxiety. For instance, trait anxiety can be correlated with increased amygdala activation coupled with higher fear expression during fear acquisition. The effects of anxiety also impact extinction learning and retention in the aspect that is impairs these processes, as well as regulating emotional responses. During or prior to fear extinction, anxious individuals portray reduced prefrontal activation and need a heightened prefrontal recruitment. Through cognitive reappraisal, there can be a successful reduction of negative emotion. There is evidence that suggests that prefrontal inhibition of the amygdala is mainly mediated by a fiber tract coming in from the ventromedial prefrontal cortex to the inhibitory cells within the amygdala. The structural integrity of this particular ventromedial prefrontal cortex-amygdala pathway is inversely proportional to trait anxiety, which suggests that compromised inhibitory function impacts emotional regulation in anxiety by heightening and impairing the process.[6] In one study, they discovered evidence of two independent dimensions of neurocognitive function that associated with trait vulnerability to anxiety.
The first entailed increased amygdala responsivity to phasic fear cues. The second involved impoverished ventral prefrontal cortical (vPFC) recruitment to downregulate both cued and contextual fear prior to omission (extinction) of the aversive unconditioned stimulus.[7]
The significance of these two aspects of neurocognitive function is that they may contribute to symptom differentiation across anxiety disorders. In this case, the mechanism of the amygdala affects developing phobic fears and the frontal mechanism impacts the maintenance of specific fears and a more generalized anxiety. It was also realized that in humans, clinical symptoms of anxiety show improved detection in sustained rather than phasic paradigms. A phasic state is when fear is adapted to the threatening stimulus – it is rapidly onset and then quickly dissipates. In a sustained state, which is an elongated state of apprehension.[8]
Information Processing Biases
According to an accumulation of research studies, there are two principal information-processing biases that are characteristic of anxiety. The first is the bias to attend toward information in relation to threats – the second is a bias towards negative interpretation of ambiguous stimuli. Neuroimaging studies also reveal that anxiety is correlated with a generation pattern of faster response times when an individual is detecting a threat stimulus or identifying a target that has been cued by this stimulus. The studies also reveal slower response times when an individual is detecting a neutral stimulus or reporting information on a neutral stimulus in the event of a threat. Anxious individuals also have the tendency to lean towards a more negative perception during a situation. For instance, they are more likely to be sensitive in a negative way towards ambiguous emotional facial expressions, homophones (interpretation of “dye/die”), and face-voice pairings. This particular type of bias to threat suggests compromised prefrontal control mechanisms that are usually engaged during attentional competition and control. High trait anxiety is also associated with increased amygdala activity towards attended and unattended stimuli, as well as decreased prefrontal activation during these times and even in the absence of a threat stimulus. These two areas (the amygdala and prefrontal cortex) may also contribute to the negative interpretation of a scenario involving anxiety. Higher trait anxiety is associated with higher amygdala BOLD responses (blood oxygenation level dependent) during passive views of neutral faces; they have a tendency to interpret these faces more negatively. Whereas healthy individuals lacking trait anxiety, the magnitude of the BOLD signals is positively correlated with the degree of negative expression interpretation, as opposed to positive. [9]
How Anxiety Impacts Decision-Making
It may be that amygdala hyperresponsitivity while evaluating, anticipating, or attending to negative stimuli may increase cognitive and effective responses to a potential threat in individuals with anxiety. On that note, any prefrontal cortex-dependent cognitive regulatory processes may be impaired during anxiety. This would reduce its potential to modulate this system. Several studies done in Catherine Hartley’s article propose that alterations in neural circuitry that involves fear learning and regulation may mediate the impact of anxiety on decision-making. Many aspects of neuroeconomics can also be taken into account. In individuals with anxiety, their cost-benefit analysis of confronting a situation or weighing risks can differ greatly from a person lacking the anxiety trait.
Ambiguity vs Risk
In another article found in the Journal of Neuroscience, a group did a functional magnetic resonance imaging experiment in order to compare ambiguity and risk. A third comparison was also added – ignorance. They had modified “learned Pavlovian CS + stimuli such that they became an ambiguous cue and contrasted evoked brain activity both with an unmodified predictive CS + (risky cue), and a cue that conveyed no information about outcome probabilities (ignorance cue)”[10]. The regions that had been shown originally engaged by decisions about ambiguous rewarding outcomes were also shown to be engaged by ambiguous outcome prediction in the sense of aversive outcomes. There was also activation found in these regions even with lack of a decision to be made. There are two forms of uncertainty that can be distinguished in the aspect of human decision-making: one is the known or calculable probability of outcomes, also known as risk – the second is ambiguity, or a scenario where there are multiple outcomes and their outcomes are completely unknown. An fMRI study was conducted to identify brain activation coding for the decision parameters of expected value (or some type of calculable outcome) and uncertainty of monetary rewards. This study showed that when a stimulus was associated with higher expected values there were monotonically increasing activations in specific areas of the striatum. When there was higher uncertainty stimulation, the imaging showed increasing activations in the lateral orbitofrontal cortex. This specific data set suggests that separate prefrontal regions are involved in risk aversion and seeking. This information was extremely useful in understanding how reward structures are setup in our own circuitry[11].
Uncertainty
There are two forms of uncertainty that can be distinguished in the aspect of human decision-making: one is the known or calculable probability of outcomes, also known as risk – the second is ambiguity, or a scenario where there are multiple outcomes and their outcomes are completely unknown. An fMRI study was conducted to identify brain activation coding for the decision parameters of expected value (or some type of calculable outcome) and uncertainty of monetary rewards. This study showed that when a stimulus was associated with higher expected values there were monotonically increasing activations in specific areas of the striatum. When there was higher uncertainty stimulation, the imaging showed increasing activations in the lateral orbitofrontal cortex. This specific data set suggests that separate prefrontal regions are involved in risk aversion and seeking. This information was extremely useful in understanding how reward structures are setup in our own circuitry.
Conclusion
The integration of many areas of the brain and their connecting circuitry mediate a large amount of emotional pathways that have a high impact on trait anxiety. The way individuals with anxiety process fear into their calculations of cost-benefit analysis likely plays a large role in their decision-making process. It has been shown that areas such as the amygdala and other areas that mediate emotions become altered in an individual who has trait anxiety. This alteration, amongst many others of similar effect, is shown to be a probable potential instigator of the outcomes that occur, resulting in anxious behaviors.
References (incase you missed some links)
- (9) Bach DR, Seymour B, and Dolan RJ. Neural Activity, Associated Passive Prediction Ambiguity, Risk for Aversive Events. J Neu. 2009;29.6: 1648-56.
- (2) Cisler JM, and Koster EHW. Mechanisms Attentional Biases Towards Threat In Anxiety Disorders: Integrative Review. Clin Psyc Rev. 2010;30.2: 203-16.
- (7) Davis M., et al. Phasic Vs Sustained Fear in Rats, Humans: Role of Extended Amygdala in Fear Vs Anxiety. Neu psyc pharm. 2010;35.1: 105-35.
- (1) Hartley, CA, and Phelps EA. Anxiety And Decision-Making. Bio Psyc. 2012;72.2: 113-118.
- (6) Indovina I., et al. Fear-Conditioning Mechanisms Assoc. with Trait Vulnerability to Anxiety in Humans. Neuron. 2011;69.3: 563-71.
- (8) Kuhnen CM, and Knutson B. The Neural Basis of Financial Risk Taking. Neuron 2005;47.5: 763-70.
- (4) Lissek S., et al. Classical Fear Conditioning in Anxiety Disorders: A Meta-Analysis. Behav Res, Ther. 2005;43.11: 1391-424.
- (3) Quirk GJ, and Mueller D. Neural Mechanisms Extinction Learning, Retrieval. Neu psyc pharm. 2008;33.1: 56-72.
- (5) Reiss S., et al. Anxiety Sensitivity, Anxiety Frequency and Prediction of Fearfulness. Behav Res, Ther. 1986;24.1: 1-8.
- (10) Tobler PN, et al. Reward Value Coding Distinct from Risk Attitude-Related Uncertainty Coding in Human Reward Systems. J Neu phys. 2007;97.2: 1621-32.
Just pointing out a grammar mistake – “girlfriends” in the second line is trying to be possessive but is missing an apostrophe. Thanks and nice article.
Fixed, thanks.